
Abstract
Cellulose synthase catalytic subunits (CESAs) play important roles in plant growth, development and disease resistance. Previous studies have shown an essential role of Arabidopsis thaliana CESA3 in plant growth. However, little is known about the role of CESA3 in species other than A. thaliana. To gain a better understanding of CESA3, the petunia (Petunia hybrida) PhCESA3 gene was isolated, and the role of PhCESA3 in plant growth was analyzed in a wide range of plants. PhCESA3 mRNA was present at varying levels in tissues examined. VIGS-mediated PhCESA3 silencing resulted in dwarfing of plant height, which was consistent with the phenotype of the A. thaliana rsw1 mutant (a temperature-sensitive allele of AtCESA1), the A. thaliana cev1 mutant (the AtCESA3 mild mutant), and the antisense AtCESA3 line. However, PhCESA3 silencing led to swollen stems, pedicels, filaments, styles and epidermal hairs as well as thickened leaves and corollas, which were not observed in the A. thaliana cev1 mutant, the rsw1 mutant and the antisense AtCESA3 line. Further micrographs showed that PhCESA3 silencing reduced the length and increased the width of cells, suggesting that PhCESA3 silencing inhibits elongation and stimulates radial expansion in petunia.
Similar content being viewed by others


Introduction
The morphology of plant organs is highly regulated through cell expansion and cell division1. The final shape and size of cells are often determined by the magnitude and direction of primary cell wall extension2,3,4. After expansion is arrested, a thick, secondary cell wall is often deposited within the boundary of the primary cell wall5.
In the past decades, significant insight into the molecular details of cellulose biosynthesis has been gained using forward and reverse genetic analyses. The plasma membrane rosettes contain the cellulose synthase catalytic subunit (CESA) proteins6,7,8,9,10. There are multiple CESA genes in the CESA gene family in plant genomes. For example, the Arabidopsis thaliana genome contains 10 CESA genes. Among these genes, CESA1, CESA3, and CESA6 are required for cellulose biosynthesis in primary cell walls11, whereas CESA4, CESA7, and CESA8 are required for cellulose biosynthesis during secondary wall deposition6,7,8,9.
In A. thaliana, the CESA3 mutation (cev1) or antisense lines have altered plant growth, including plant height, leaf size and petal size, as well as altered fertility12,13,14, which are the important ornamental characteristics for ornamental plants. In addition, CESA3 is involved in resistance, activating hormone signaling and cellulose microfibril crystallinity10,12,14,15,16. Studies on A. thaliana and Nicotiana tabacum L. variety Samsun NN have demonstrated that the expression of the CESA3 cellulose synthase gene that contains a point mutation, named ixr1–2, results in greater conversion of plant-derived cellulose to fermentable sugars17.
Petunia (Petunia hybrida) is often used as a model for studying ornamental plant growth and development. A developmental map of petunia petals has been assembled by reconstructing the pattern of cell expansion through measurement of cell size18.
Here, we isolated the full-length cDNA of PhCESA3, and its spatiotemporal expression was analyzed. VIGS-mediated PhCESA3 silencing resulted in short but swollen stems, pedicels, filaments, styles and epidermal hairs as well as small and thickened leaves and petals. Further micrographs showed a reduced length and increased width of cells in PhCESA3-silenced plants. These results showed the important role of PhCESA3 in cell elongation and radial expansion. In addition, fertility was reduced in PhCESA3-silenced plants.
Results
Isolation of P. hybrida PhCESA3 cDNA
The full-length cDNA of PhCESA3 was isolated from a cDNA library prepared from the corollas of petunia by homologous cloning and the RACE-PCR method. PhCESA3 was predicted to encode a protein of 1083 amino acids with a calculated molecular weight of 120.8 kDa and a pI of 7.02.
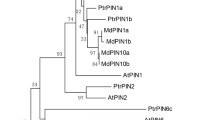
The multiple sequence alignments of PhCESA3 protein in the petunia and A. thaliana are presented in Fig. S1. PhCESA3 shares 86.5% amino acid sequence identity with AtCESA3 (Table S1). In addition to a putative zinc-binding domain (at amino acid residues 31–76) and two putative transmembrane helices in the N-terminus, PhCESA3 contains six putative transmembrane helices in the C-terminus (Fig. S1) similar to A. thaliana CESA proteins, and the C-terminus is strikingly conserved19,20. In the course of the publication of the study, the petunia genome sequences were released21 (https://www.sgn.cornell.edu/). By blasting against Sol Genomics Network with A. thaliana AtCESAs, nine PhCESAs, including PhCESA3, were obtained in the petunia genome. The PhCESAs are named PhCESA1, PhCESA2A, PhCESA2B, PhCESA2C, PhCESA3, PhCESA4, PhCESA6, PhCESA7 and PhCESA8 based on the identity of their encoding amino acid sequences with the A. thaliana AtCESAs (Table S1). The phylogenetic tree of the PhCESAs and A. thaliana AtCESAs shown in Fig. 1 further reveals that the cloned PhCESA3 is the ortholog of AtCESA3.

Petunia PhCESAs were aligned with Arabidopsis thaliana AtCESA1 (AT4G32410), AtCESA2 (AT4G39350), AtCESA3 (AT5G05170), AtCESA4 (AT5G44030), AtCESA5 (AT5G09870), AtCESA6 (AT5G64740), AtCESA7 (AT5G17420), AtCESA8 (AT4G18780), AtCESA9 (AT2G21770), and AtCESA10 (AT2G25540). Petunia PhCESAs come from Sol Genomics Network. PhCESA1, Peaxi162Scf00073g01124; PhCESA2A, Peaxi162Scf00025g00276; PhCESA2C, Peaxi162Scf00079g02433; PhCESA2B, Peaxi162Scf01294g00032; PhCESA3, Peaxi162Scf00953g00116; PhCESA4, Peaxi162Scf00415g00078; PhCESA6, Peaxi162Scf00041g00017; PhCESA7, Peaxi162Scf00401g00329; PhCESA8, Peaxi162Scf00371g00318.
Expression patterns of PhCESA3 mRNA
As a step toward functional analysis, we examined the spatiotemporal expression of PhCESA3 petunia ‘Ultra’ using quantitative real-time PCR (qPCR) with gene-specific primers, and actin was used as an internal control. PhCESA3 transcripts were present in root, stem, leaf, and corolla, with varying patterns of expression (Fig. 2). PhCESA3 mRNA was slightly more abundant in roots and stems compared to corollas.

R, roots; L, leaves; S, stems; C, corollas. Relative expression levels are shown as fold change values. Data are presented as the mean ± SD (n = 3).
VIGS-mediated PhCESA3 silencing leads to plant dwarfing
To explore the function of PhCESA3, we generated a TRV-PhCESA3 vector that contains a 266 bp fragment of the 3′ untranslated sequence of PhCESA3 cDNA to test the silencing of PhCESA3 by virus-induced gene silencing (VIGS)22. Five weeks after TRV infection, plants infected with TRV-PhCESA3 showed a significant dwarfing phenotype. The stems and internodes were shorter and wider than those of the empty TRV-infected control plants (Fig. 3a–e; Table 1). The silenced stems were about half the length of the control. The diameter of silenced stems was significantly increased. In addition, there were shorter stem epidermal hairs on PhCESA3-silenced plants compared to control plants (Fig. 3f). PhCESA3 silencing also resulted in smaller and thicker leaves than those of control plants (Fig. 3g and Table 1). Analysis by qPCR showed decreased PhCESA3 mRNA levels in corollas, leaves and stems in plants treated with TRV-PhCESA3 compared to those in control plants (Figs 4 and S2 and 3).

(a,b) Five-week-old VIGS-mediated PhCESA3-silenced plants (middle and right) compared to control plants (left). (c,d) Stem inflorescence of PhCESA3-silenced plants (right) compared with that of control plants (left) with leaves (c) and without leaves (d). (e,f) Internodes (e) and hairs (f) of stems of PhCESA3-silenced plants (right) compared with those of control plants (left). (g–i,m), Mature leaves (g), buds (h), opened flowers (i) and stigmas (m) of PhCESA3-silenced plants (bottom) compared with those of control plants (top). (j–l,n–q), Pedicel (j), filament (k), style (l), filament surface (n), style surface (o), ovary (unpollinated) (p) and young fruit (q) of PhCESA3-silenced plants (right) compared with those of control plants (left).

Relative expression levels are shown as fold change values. Data are presented as the mean ± SD (n = 3). Statistical analysis was performed using Student t test with 3 replicates. Asterisk means significant difference at P = 0. 05 level.
In addition, PhCESA3 silencing has no significantly effect on the expression of the other 8 PhCESAs in corollas (Fig. 4), leaves (Fig. S2) and stems (Fig. S3). PhCESA2B, PhCESA2C, and PhCESA6 mRNA levels were not detected by qPCR in either TRV-PhCESA3-treated or control corollas and PhCESA6 mRNA levels were not also detected in either TRV-PhCESA3-treated or control leaves.
PhCESA3 silencing alters flower organ growth
PhCESA3 silencing reduced the height of stems but did not change the flowering time. However, altered flower organ growth was observed in PhCESA3-silenced plants. PhCESA3 silencing reduced the size but increased the thickness of petal limbs, and it reduced the height of petal tubes (Fig. 3h and i; Table 1).
The pedicels (fruit stalks), filaments and styles were shorter and wider in PhCESA3-silenced plants than those in control plants (Fig. 3j–l; Table 1). The lengths of pedicels (fruit stalks), styles, and filaments in PhCESA3-silenced plants were about two-thirds, half and half of those of control plants, respectively (Table 1). The diameters of pedicels and styles of PhCESA3-silenced plants were about one and half, and two fold of those of control plants, respectively (Table 1). In addition, the styles and filaments were semi-transparent and loose, and their surface was rough in PhCESA3-silenced plants (Fig. 3n and o). In PhCESA3-silenced plants, the stigma was small and deformed (Fig. 3m), and the unpollinated ovaries and young fruits were often deformed (Fig. 3p and q).
PhCESA3 silencing leads to a significant reduction in cellulose content
To further characterize the function of PhCESA3, cellulose content was analyzed in plants treated with TRV and TRV-PhCESA3 treatment plant. PhCESA3-silenced plants showed a significant decrease in total cellulose content of the stems and leaves (Fig. 5a and b). In 5-week-old plants, mature stems in PhCESA3-silenced plants had about 80% and mature leaves had about 70% of the control level of cellulose (Fig. 5a and b).

Cellulose was measured in stem segments (a) and leaves (b) from wild-type and PhCESA3-silenced plants. Standard error bars are shown (n = 6). Statistical analysis was performed using Student t test with 6 replicates. Asterisk means significant difference at P = 0.05 level.
PhCESA3 silencing leads to a significant reduction in fertility
The effects of PhCESA3 silencing on fertility were examined. PhCESA3 silencing caused a significant reduction in fertility, resulting in fewer than 100 seeds per fruit. More seeds per fruit were obtained in reciprocal crosses between PhCESA3-silenced plants and control plants when PhCESA3-silenced plants were used as the pollen donor or receptor (Table 2) but fewer than crosses between both control plants as parents, indicating reduced female and male fertility. Reciprocal crosses with control plants showed that the female reproductive capacity was more strongly affected than male reproductive capacity in PhCESA3-silenced plants (Table 2).
PhCESA3 silencing inhibits cell elongation and causes cell swelling
Because PhCESA3 silencing altered the growth of stems, leaves and flowers, we further examined these organs in detail. Scanning electron micrographs showed reduced cell length and increased cell width of the epidermal cells of stems, pedicels, filaments and styles in PhCESA3-silenced plants (Fig. 6a–d; Table 3). Optical micrographs of the epidermis of stems and pedicels showed similar results (Fig. S4a and b).

(a–d), Epidermal cells of stem (a), pedicel (b), filament (c) and style (d) showing reduced cell length and increased cell width in PhCESA3-silenced plants (right, bottom in (d)) compared to control plants (left, top in (d)). (e,f), Adaxial (e) and abaxial (f) epidermal cells of leaves showing the reduced cell size in PhCESA3-silenced plants (right) compared to control plants (left). (g,h) Adaxial (g) and abaxial (h) epidermal cells of petal limb showing the reduced cell size in PhCESA3-silenced plants (right) compared to control plants (left). (i), Pedicels showing the reduced number of trichomes in PhCESA3-silenced plants (right) compared to control plants (left). (j) Trichomes of pedicel showing the reduced trichome length and more collapsed trichomes in PhCESA3-silenced plants (right) compared to control plants (left). (k) Pollen grains showing the abnormal pollen mother cell in PhCESA3-silenced plants (right) compared to control plants (left). Bars = 50 μm in (a–f); bars = 20 μm in (g–h); bar = 1 mm in (i); bars = 200 μm in (j); bars = 10 μm in (k).
As visualized in transverse and longitudinal sections, PhCESA3 silencing also reduced the length and increased the width of the cortical cells of stems, pedicels, and filaments, but the number of cortical cells did not change (Fig. 7a,b,e,f and l; Table 3), indicating that the swollen stems were attributed to an increased cell width rather than a decrease in cell number. Hand-cut sections also showed similar results (Fig. S5a and b). The transverse sections showed wider pedicels, filaments and styles in PhCESA3-silenced plants than those in control plants (Fig. 7d,j,m; Table 1). In addition, the arrangement of cortical cells in stems, pedicels and filaments was irregular and close in PhCESA3-silenced plants compared with that of control plants (Fig. 7b,e,l). The cortical cells were often round or oval shaped in control plants, while the cortical cells were often irregular polygon shaped in PhCESA3-silenced plants (Fig. 7b,e and l).

(a) Stem longitudinal section showing the increased width of cortex and cortical cells in PhCESA3-silenced plants (right) compared to control plants (left). (b) Stem transverse section showing the uneven surface in PhCESA3-silenced plants (right) compared to control plants (left). (c) Leaf transverse section showing the thickened leaf in the PhCESA3-silenced plants (right) compared to the control plant (left). (d) Pedicel transverse section showing the thickened pedicel in the PhCESA3-silenced plants (right) compared to the control plant (left). (e,f) Pedicel transverse (e) and longitudinal (f) sections showing the increased width of cortical cells in PhCESA3-silenced plants (right) compared to control plants (left). (g) Petal limb transverse section showing the thickened petal limb in the PhCESA3-silenced plants (right) compared to the control plant (left). Solid and hollow arrows indicate the cell shape of the adaxial and abaxial sides, respectively. (h) The adaxial epidermal cells of petal limb showing sleek cells in the PhCESA3-silenced plants (bottom) compared to the control plant (top). (i) The abaxial epidermal cells of petal limb showing the uneven size and irregular shape of cells in the PhCESA3-silenced plants (bottom) compared to the control plant (top). (j,m) Transverse section showing the thickened filament (j) and style (m) in the PhCESA3-silenced plants (right) compared to the control plant (left). (k,n) Marrow transverse section of filament and style showing the irregularly arranged marrow cells and different cell sizes in the PhCESA3-silenced plants (right) compared to the control plant (left). (l) Filament cortex transverse section showing the irregularly arranged cortical cells and uneven cell sizes in the PhCESA3-silenced plants (right) compared to the control plant (left). (o), Style transverse section showing the width of epidermal cells in the PhCESA3-silenced plants (right) compared to the wild-type plant (left). The cortex (co), cambium (ca), and xylem elements (xe) are indicated. Bars = 250 μm in (a,b,f,j,m); bars = 100 μm in (e,c); bars = 500 μm in (d,l); bars = 50 μm in (g,k,n,o); bars = 10 μm in (h,i).
Scanning electron micrographs of adaxial and abaxial epidermal cells of leaves and petal limbs showed reduced cell size in PhCESA3-silenced plants (Fig. 6e and h). The reduced proportion of leaves and petal limbs was similar to that of the epidermal cells of leaves and petal limbs. Optical micrographs of the epidermis of leaves and petals showed similar results (Fig. S4c–f). As visualized in transverse sections, PhCESA3 silencing increased the width of the mesophyll cells of leaves and petals (Fig. 7c and g), while the number of mesophyll cells did not change, indicating that the thickened leaves and petals were attributed to an increased cell size rather than cell number.
In addition, the sections of petal limbs revealed that the outside shape of the epidermal cells of the adaxial side of petal limbs from control plants was often sharp, while the outside shape of the same cells from PhCESA3-silenced plants was curved (Fig. 7g and h). The epidermal cells of the abaxial side of petal limbs of control plants showed a similar size and good arrangement, but the same cells in PhCESA3-silenced plants had different sizes, different shapes, and irregular arrangement (Fig. 7g and i). Transmission electron micrographs of cells from petal limbs confirmed these results (Fig. 8e–g).

Mature inflorescence stems were cross-sectioned for imaging of interfascicular fiber cell walls. (a) Stem epidermal cells showing the reduced thickness of cell wall in the PhCESA3-silenced plants (right) compared to the control plant (left). (b) Stem cortical cells showing similar cell wall thickness in PhCESA3-silenced plants (right) and wild-type plants (left). (c) Chloroplast of stem cortical cells showing the reduced starch granules in the PhCESA3-silenced plants (right) compared to control plants (left). (d) Stem xylem cells showing similar cell wall thickness in PhCESA3-silenced plants (right) and control plants (left). (e) Petal limb epidermal cells showing the reduced cell wall thickness in PhCESA3-silenced plants (right) compared to control plants (left). (f) Petal limb upper epidermal cells showing the change of cell shape in PhCESA3-silenced plants (bottom) compared to control plants (top). (g) Petal limb lower epidermal cells showing the change of cell shape and reduced cell wall thickness in PhCESA3-silenced plants (right) compared to control plants (left). (h) Filament epidermal cells showing the change of cell shape, cell size and reduced cell wall thickness in PhCESA3-silenced plants (right) compared to control plants (left). (i) Filament cortical cells showing changed cell shape and size but similar cell wall thickness in PhCESA3-silenced plants (right) compared to control plants (left). Bars = 10 μm in (a,b,f–i); bars = 1 μm in (c); bars = 2.5 μm in (d); bars = 5 μm in (e).
Scanning electron micrographs showed that the epidermal cells of petals in PhCESA3-silenced plants were often collapsed while those of control plants were plump (Fig. 6g). The collapsed cells may have been due to thin and fragile epidermal cell walls in PhCESA3-silenced plants compared to those of control plants as revealed by transmission electron microscopy (Fig. 8e).
The transverse sections of filaments and styles showed the wide marrow in PhCESA3-silenced plants compared with that of the control (Fig. 7k and n). Moreover, the myeloid cells of control plants were similar in size and had a good arrangement while those of PhCESA3-silenced plants had different sizes, different shapes and irregular arrangement (Fig. 7k and n). In addition, wider epidermal cells of style were observed in the PhCESA3-silenced plants compared to those in the control plants (Fig. 7o).
Scanning electron and optical micrographs of hand-cut sections and stereomicroscope micrographs all showed that PhCESA3 silencing reduced the length and increased the width of cells of epidermal hairs in stems and pedicels (Fig. S5c and d). Scanning electron microscopy showed the increase of number of epidermal hairs of pedicels in the PhCESA3-silenced plants (Fig. 6i). In addition, scanning electron micrographs showed collapsed epidermal hairs on PhCESA3-silenced plants, which may be attributed to the thin and fragile cell walls in PhCESA3-silenced plants (Fig. 6j).
In addition, scanning electron micrographs of pollen grains showed that parts of the pollen cell wall were collapsed in PhCESA3-silenced plants, but the pollen cells in control plants were plump (Fig. 6k), which may partially explain the sterility of PhCESA3-silenced plants.
PhCESA3 silencing alters the ultrastructure of cells
To investigate if PhCESA3 silencing affects the ultrastructure of cells, we examined the thickness of cell walls. Transmission electron micrographs of cell walls revealed that the epidermal cell walls of stems, petal limbs and filaments in PhCESA3-silenced plants were thin compared to those of control plants (Fig. 8a,e,f,g and h; Table 4), while the thickness of the cell walls of cortical cells and xylem cells of stems was not significantly different between the plants (Fig. 8b and d; Table 4). In addition, we observed that the starch granules in chloroplasts of stem cortical cells in PhCESA3-silenced plants were rare compared to control plants (Fig. 8c).
Discussion
CESA3, which encodes the catalytic subunit 3 of cellulose synthase, has been shown to be involved in the biosynthesis of primary cell walls in A. thaliana12,13,14,15. We isolated a petunia homolog of the A. thaliana CESA3 gene (PhCESA3), and we analyzed the spatial and temporal regulation of PhCESA3 expression. Furthermore, we produced VIGS-mediated PhCESA3-silenced plants and compared these plants to control plants to examine the role of PhCESA3 in plant growth and development.
The predicted peptide sequence of PhCESA3 shows high similarity to A. thaliana CESAs throughout the protein with a putative zinc-binding domain and two putative transmembrane helices in the N-terminus as well as six putative transmembrane helices in the C-terminus (Fig. S1). PhCESA3 has the homology with AtCESA3 in A. thaliana, which indicates that PhCESA3 is the homolog of AtCESA3. PhCESA3 mRNA was detected in all organs examined but was present at different levels in different organs (Fig. 2), indicating that PhCESA3 expression is spatially regulated.
In the flowers of plants treated with TRV-PhCESA3, PhCESA3 mRNA levels showed decreased compared to those of control flowers, while the mRNA levels of other PhCESAs were not significantly changed compared to those of control flowers (Fig. 4), which showed the specific silencing of PhCESA3 in plants treated with TRV-PhCESA3.
In A. thaliana, mutants of AtCESA4, 7, and 8 specifically show a cellulose defect in the secondary wall of the xylem15,23,24, whereas AtCESA1, 3 and 6 are mainly involved in cellulose synthesis in the primary cell wall5,25,26. AtCESA1 and AtCESA3 are required for cellulose synthesis in the primary cell wall, whereas AtCESA2 and AtCESA6 may be at least partially redundant9. Null alleles of AtCESA3 and AtCESA1 are embryo lethal27,28. In A. thaliana, the rsw1 mutant (the temperature-sensitive allele of AtCESA1) the antisense AtCESA3 line, and the cev1 mutant (a mild mutation of AtCESA3) results in dwarfing of plants, small leaves, small petals, short pedicels, short filaments, short styles, and reduced fertility compared to control plants12,13,14,15. The short stamen filament surface of the rsw1 mutant is crumpled29. In tobacco, using the VIGS-method silencing of NtCESA1, the homolog of AtCESA3, leads to shorter internode lengths, small leaves, and a dwarf phenotype30. In this study, similar results were obtained in the PhCESA3-silenced plants (Fig. 3a–e,g,i–l). Both the antisense AtCESA3 line and rsw1 mutant affect female reproductive competence more severely than male competence in A. thaliana13,29, which is consistent with the results in this study (Table 2). However, PhCESA3 silencing caused swollen stems, pedicels, filaments, styles and epidermal hairs, which were not observed in the AtCESA3 antisense line, the cev1 mutant and the rsw1 mutant in A. thaliana12,13,14,15,29. Further, scanning, transmission and optical micrographs showed that PhCESA3 silencing caused short and swollen cells of these organs or tissues, which was consistent with the corresponding phenotypes. Thus, PhCESA3 silencing resulted in primary cell wall extension in length and width. Importantly, growing the rsw1 mutant at its restrictive temperature inhibits root and hypocotyl elongation as well as promotes radial swelling29. Mutations in A. thaliana AtCESA3 and AtCESA6 confer resistance to isoxaben (an herbicide that specifically inhibits cellulose) and isoxaben, which causes radial swelling of roots15. Taken together, with strong inhibition of cellulose synthesis, whether genetic or chemical (i.e., with herbicides such as isoxaben), organ elongation is inhibited and radial expansion is stimulated. However, the organs that exhibit radial expansion induced by CESA silencing are different in various species.
Among the ten AtCESAs in A. thaliana, AtCESA3 (IXR1/CEV1) is suggested to be essential for depositing cellulose in primary walls10. When changes to primary walls alter cell expansion and/or cell divisions, changes to cell shape, growth, and morphogenesis occur. Changes to thickening after growth stops occur too late to affect these processes, resulting in normal morphology but changes in mechanical properties13. Similar to the A. thaliana AtCESA3 mutant, PhCESA3 silencing resulted in significant changes in the cell size and shape in several organs. Moreover, the cellulose content of mature stems and leaves from PhCESA3-silenced plants was significantly decreased compared to that of control plants. These results suggested that PhCESA3 mainly functions for catalyzing the biosynthesis of cellulose deposited to the primary wall. Ultrastructural studies clearly showed that the cell walls of the epidermis in stems and petals, which are heavily thickened during primary growth31, were thinner, thus further supporting the involvement of PhCESA3 in depositing cellulose in the primary wall. The thickness of the cell walls of cortical cells and xylem cells of stems did not significantly change (Fig. 8b and d; Table 4), suggesting that PhCESA3 is not involved in depositing cellulose in thickening walls after cell growth stops.
In this study, the shorter stems, pedicels, filaments and styles in PhCESA3-silenced plants were attributed to reduction in cell length rather than cell number, and these swollen organs in PhCESA3-silenced plants were attributed to increased cell width rather than cell number (Fig. 6a–d; Fig. 7a,b,e,f,j and l). These results implicated that PhCESA3 is mainly involved in cell elongation rather than cell division, which is consistent with results obtained with the antisense AtCESA3 line in A. thaliana12,13,14,15. In addition, plant organ shaping requires control over cell wall expansion anisotropy, which is characterized by the direction and degree of anisotropy32. In this study, the inhibited elongation and stimulated radial expansion of several organs in PhCESA3-silenced plants showed the change of the direction and degree of anisotropy of cell wall.
Here, PhCESA3 silencing resulted in changes of plant height, flower size and fertility, which are the important features of ornamental plants. These results suggested the important role of PhCESA3 in controlling plant stature and form. By using specific promoters, changes in CESA3 expression in different organs may create an excellent variety for ornamental plants. Therefore, PhCESA3 may be utilized in petunia breeding for changing the plant height, flower size and fertility.
Methods
Plant material
Petunia ‘Ultra’ plants were grown under greenhouse conditions (22–25 °C, 14 h light/10 h dark). Flowers were emasculated 1 d before the flowers were fully open to prevent self-pollination. Eight to ten petunia flowers were harvested at anthesis (corollas 90° reflexed) stages and were then placed immediately in tap water. Stem, leaves and roots were collected from plants at the vegetative stage when the plants were approximately 10 cm in height. All tissues were frozen in liquid nitrogen and stored at −80 °C until used for RNA extraction. Fresh weights were measured immediately before freezing. All experiments were conducted at least three times with independently collected and extracted tissues unless otherwise noted.
RNA extraction, RT-PCR and cloning of the petunia PhCESA3 gene
Total RNA was extracted and reverse-transcribed according to the methods of Liu et al.33. The partial sequences of PhCESA3 were obtained using a computational identification approach34. Briefly, TBLASTN analysis against the GenBank EST database (http://www.ncbi.nlm.nih.gov) with A. thaliana CESA3 identified 1 petunia clone, FN016492, which encodes putative proteins that displayed conservation with A. thaliana CESA3. The remaining 5′ and 3′ cDNA sequences of PhCESA3 were isolated by RACE using a specific primer (Table S2), and its full-length cDNA was isolated by RT-PCR.
Sequence analysis
Alignments were performed, and a phylogenetic tree was generated using the DNAMAN software. Identity search for nucleotides and translated amino acids was performed using the National Center for Biotechnology Information (NCBI) BLAST network server (http://www.ncbi.nlm.gov/BLAST).
Quantitative real-time PCR assays
Quantitative real-time PCR (qPCR) assays were performed according to previous methods33. Analyses were conducted following the Minimum Information for Publication of Quantitative Real-Time PCR Experiments guidelines35. Petunia Actin (accession no. FN014209) and Cyclophilin (CYP) (accession no. EST883944) genes were used as the internal reference genes to quantify the cDNA abundance36. Similar results were obtained for both reference genes. The data presented in the body of the text represent relative expression values calculated using Actin, and the data presented in the attachment represent relative expression values calculated using CYP. The sequences of all primers used for qPCR analysis are described in Table S3.
Agroinoculation of TRV vectors
To generate pTRV2 containing the 3′ untranslated region of PhCESA3 (TRV2-PhCESA3), the gene sequence of 266 bp was PCR amplified using forward primers and reverse primers (Table S4), and the PCR products were inserted into the pTRV2 vector. Agrobacterium tumefaciens (strain GV3101) transformed with pTRV1 and pTRV2 derivatives were prepared as previously described22,37. The Agrobacterium cells grown overnight were harvested and resuspended in inoculation buffer containing 10 mM MES, 200 mM acetosyringone, and 10 mM MgCl2 to an OD550 of 10. Following an additional 3 h of incubation at 28 °C, bacteria transformed with pTRV1 were mixed with bacteria containing the pTRV2 derivatives in a 1:1 ratio, and 200 to 400 μL of this mixture was injected into the stem or applied on the cut surface after removing the apical meristems of petunia plantlets. Approximately thirty plants were vaccinated with each vector. The inoculated plants were grown under greenhouse conditions (22–25 °C, 14 h light/10 h dark).
Cellulose measurement
Cellulose content in mature stems and leaves was measured by a previously published method38,39. Analysis was conducted with material from a single plant using three to five fully expanded leaves (from 5-week-old plants) or a 3 cm segment from the base of the primary inflorescence stem. Stems were chopped with a razor blade, and a crude cell wall fraction was obtained by extracting the soluble material with two changes of 70% ethanol at 70 °C for 1 h each40. The ethanol was removed, and the samples were dried under vacuum. The dry weight of the wall material was recorded. Cellulose was measured according to a previously published method39.
Scanning electron micrographs
The stems, leaves, pedicels, petal limbs, filaments, styles and anthers from PhCESA3-silenced plants and control plants were cut into 3–5 mm2 pieces. The samples were fixed in 4% glutaraldehyde in 0.1 mol/L PBS (pH 7.2) for 4 h at 4 °C and then washed 3 times in the same buffer, which was followed by post-fixation in 1% osmium tetroxide for 2 h at room temperature and 3 rinses using the same buffer. Samples were dehydrated in increasing grades of ethanol and then dried with a critical point drier (CPD 030, Switzerland, Bal-Tec). The dried samples were fixed on the sample stage and coated with gold by ion sputtering equipment. Samples were observed with a scanning electron microscope (XL-30-ESEM, The Netherlands, FEI) at 10 kV acceleration and photographed.
Paraffin sections
The stems, leaves, pedicels and styles from plants were cut into 5 mm × 5 mm × 5 mm pieces. The samples were fixed in FAA fixative solution (every 100 ml of FAA fixative solution contains 90 ml of 50% or 70% ethanol, 5 ml of acetic acid, and of 5 ml formalin) for 24 h at room temperature and then washed in running water for 24 h. Samples were stained by hematoxylin for 4 d and then washed in running water for 24 h, which was followed by dehydration in increasing grades of ethanol. A graded chloroform series was used for clearing, and the samples were embedded in paraffin. Paraffin sections were cut to a thickness of 8 μm on a Leica RM2235 followed by dewaxing with xylene. Lastly, slides were sealed with neutral resin. Sections were observed and photographed with a Zeiss Scope.A1 microscope.
Semi-thin sections
The pedicels, petal limbs and filaments from PhCESA3-silenced plants and control plants were cut into 1 mm × 1 mm × 0.5 mm pieces. The specimens were fixed in 2.5% paraformaldehyde/3.0% glutaraldehyde in 0.1 mol/L PBS (pH 7.2) for 4 h at 4 °C, and the specimens were then washed 3 times in the same buffer, which was followed by post-fixation in 1% osmium tetroxide for 2 h at room temperature and 3 rinses using the same buffer. The specimens were dehydrated in a graded ethanol series and embedded in Epon812 (SPI Supplies Division of Structure Probe Inc., West Chester, PA, USA). Polymerization took place for 24 h at 40 °C, which was followed by 24 h at 60 °C. Specimens were cut to a thickness of 1 μm on a Leica RM2155 and were stained with 0.5% toluidine blue. Sections were observed and photographed with a Leica DMLB microscope.
Transmission electron micrograph
The stems, petal limbs and filaments from PhCESA3-silenced plants and control plants were cut into <1 mm3 pieces. The samples were fixed in 4% glutaraldehyde in 0.1 mol/L PBS (pH 7.2) for 4 h at 4 °C and then washed 3 times in the same buffer, which was followed by post-fixation in 1% osmium tetroxide for 2 h at room temperature and 3 rinses using the same buffer. Samples were dehydrated in increasing grades of ethanol and embedded in Epon812 (containing 51.64% Epon812, 5.37% DDSA, 42.99% MNA and 1.5% DMP-30). Polymerization took place for 24 h at 45 °C, which was followed by 24 h at 60 °C. Ultrathin sections (100 nm thick) were cut with a Leica ULTACUT ultramicrotome and deposited on a 200 mesh copper net with support film. Samples were stained with a 2% uranyl acetate solution followed by a 6% lead citrate solution, and samples were visualized with a Tecnai 12 transmission electron microscope (FEI, Eindhoven, The Netherlands) at 80 kV acceleration and photographed.
Additional Information
How to cite this article: Yang, W. et al. PhCESA3 silencing inhibits elongation and stimulates radial expansion in petunia. Sci. Rep. 7, 41471; doi: 10.1038/srep41471 (2017).
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
Meyerowitz, E. M. Genetic control of cell division patterns in developing plants. Cell 88, 299–308 (1997).
Martin, C., Bhatt, K. & Baumann, K. Shaping in plant cells. Curr Opin Plant Biol. 4, 540–549 (2001).
Kotilainen, M. et al. GEG participates in the regulation of cell and organ shape during corolla and carpel development in Gerbera hybrida. Plant Cell 11, 1093–1104 (1999).
Smith, L. G. Cytoskeletal control of plant cell shape: getting the fine points. Curr Opin Plant Biol. 6, 63–73 (2003).
Fagard, M. et al. PROCUSTE1 encodes a cellulose synthase required for normal cell elongation specifically in roots and dark-grown hypocotyls of Arabidopsis. Plant Cell 12, 2409–2423 (2000).
Taylor, N. G., Gardiner, J. C., Whiteman, R. & Turner, S. R. Cellulose synthesis in the Arabidopsis secondary cell wall. Cellulose 11, 329–338 (2004).
Doblin, M. S., Kurek, I., Jacob-Wilk, D. & Delmer, D. P. Cellulose biosynthesis in plants: from genes to rosettes. Plant Cell Physiol. 43, 1407–1420 (2002).
Scheible, W. & Pauly, M. Glycosyltransferases and cell wall biosynthesis: novel players and insights. Curr Opin Plant Biol. 7, 285–295 (2004).
Somerville, C. Cellulose synthesis in higher plants. Annu. Rev. Cell Dev. Biol. 22, 53–78 (2006).
Doblin, M. S., Kurek, I., Jacob-Wilk, D. & Delmer, D. P. Cellulose biosynthesis in plants: from genes to rosettes. Plant Cell Physiol. 43, 1407–1420 (2002).
Robert, S., Mouille, G. & Höfte, H. The mechanism and regulation of cellulose synthesis in primary walls: lessons from cellulose-deficient Arabidopsis mutants. Cellulose. 11, 351–364 (2004).
Ellis, C. & Turner, J. G. The Arabidopsis mutant cev1 has constitutively active jasmonate and ethylene signal pathways and enhanced resistance to pathogens. Plant Cell. 13, 1025–1033 (2001).
Burn, J. E., Hocart, C. H., Birch, R. J., Cork, A. C. & Williamson, R. E. Functional Analysis of the Cellulose Synthase Genes CesA1, CesA2, and CesA3 in Arabidopsis. Plant Physiol. 129, 797–807 (2002).
Ellis, C., Karafyllidis, I., Wasternack, C. & Turner, J. G. The Arabidopsis mutant cev1 links cell wall signaling to jasmonate and ethylene responses. Plant Cell 14, 1557–1566 (2002).
Scheible, W., Eshed, R., Richmond, T., Delmer, D. & Somerville, C. Modifications of cellulose synthase confer resistance to isoxaben and thiazolidinone herbicides in Arabidopsis Ixr1 mutants. P Natl Acad Sci USA 98, 10079–10084 (2001).
Harris, D. M. et al. Cellulose microfibril crystallinity is reduced by mutating C-terminal transmembrane region residues CESA1A903V and CESA3T942I of cellulose synthase. P Natl Acad Sci USA 109, 4098–4103 (2012).
Sahoo, D. & Maiti, I. Biomass derived from transgenic tobacco expressing the Arabidopsis CESA3 ixr1–2 gene exhibits improved saccharification. Acta Biol Hung. 65, 189–204 (2014).
Reale, L. et al. Patterns of cell division and expansion in developing petals of Petunia hybrida . Sexual Plant Reproduction 15, 123–132 (2002).
Holland, N. et al. A comparative analysis of the plant cellulose synthase (CesA) gene family. Plant Physiol. 123, 1313–1324 (2000).
Chen, S., Ehrhardt, D. W. & Somerville, C. R. Mutations of cellulose synthase (CESA1) phosphorylation sites modulate anisotropic cell expansion and bidirectional mobility of cellulose synthase. P Natl Acad Sci USA 107, 17188–17193 (2010).
Bombarely, A. et al. Insight into the evolution of the Solanaceae from the parental genomes of Petunia hybrida . Nature plants 2, 16074 (2016).
Tan, Y., Liu, J., Huang, F., Zhong, S. & Yu, Y. PhGRL2 protein, interacting with PhACO1, is involved in flower senescence in the petunia. Mol Plant 7, 1384–1387 (2014).
Taylor, N. G., Laurie, S. & Turner, S. R. Multiple cellulose synthase catalytic subunits are required for cellulose synthesis in Arabidopsis. Plant Cell 12, 2529–2539 (2000).
Taylor, N. G., Howells, R. M., Huttly, A. K., Vickers, K. & Turner, S. R. Interactions among three distinct CesA proteins essential for cellulose synthesis. P Natl Acad Sci USA 100, 1450–1455 (2003).
Arioli, T. et al. Molecular analysis of cellulose biosynthesis in Arabidopsis. Science 279, 717–720 (1998).
Richmond, T. Higher plant cellulose synthases. Genome Biol. 1, 1 (2000).
Beeckman, T. et al. Genetic complexity of cellulose synthase A gene function in Arabidopsis embryogenesis. Plant Physiol. 130, 1883–1893 (2002).
Gillmor, C. S., Poindexter, P., Lorieau, J., Palcic, M. M. & Somerville, C. α-Glucosidase I is required for cellulose biosynthesis and morphogenesis in Arabidopsis. The Journal of Cell Biology 156, 1003–1013 (2002).
Williamson, R. E. et al. Morphology ofrsw1, a cellulose-deficient mutant of Arabidopsis thaliana . Protoplasma 215, 116–127 (2001).
Burton, R. A. et al. Virus-induced silencing of a plant cellulose synthase gene. Plant Cell 12, 691–705 (2000).
Engels, F. M. & Jung, H. G. Alfalfa stem tissues: cell-wall development and lignification. Ann Bot-London 82, 561–568 (1998).
Baskin, T. I. Anisotropic expansion of the plant cell wall. Annu Rev Cell Dev Bi. 21, 203 (2005).
Liu, J. et al. Identification and expression analysis of ERF transcription factor genes in petunia during flower senescence and in response to hormone treatments. J Exp Bot 62, 825–840 (2011).
Yu, Y., Wang, H., Liu, J., Fu, Z. & Wang, J. Transcriptional regulation of two RTE‐like genes of carnation during flower senescence and upon ethylene exposure, wounding treatment and sucrose supply. Plant Biology 13, 719–724 (2011).
Bustin, S. A. et al. The MIQE guidelines: minimum information for publication of quantitative real-time PCR experiments. Clin Chem. 55, 611–622 (2009).
Mallona, I., Lischewski, S., Weiss, J., Hause, B. & Egea-Cortines, M. Validation of reference genes for quantitative real-time PCR during leaf and flower development in Petunia hybrida. BMC Plant Biol. 10, 4 (2010).
Spitzer-Rimon, B. et al. EOBII, a gene encoding a flower-specific regulator of phenylpropanoid volatiles’ biosynthesis in petunia. Plant Cell 22, 1961–1976 (2010).
Potikha, T. & Delmer, D. P. A mutant of Arabidopsis thaliana displaying altered patterns of cellulose deposition. Plant J. 7, 453–460 (1995).
Updegraff, D. M. Semimicro determination of cellulose inbiological materials. Anal Biochem. 32, 420–424 (1969).
Reiter, W., Chapple, C. C. & Somerville, C. R. Altered growth and cell walls in a fucose-deficient mutant of Arabidopsis. Science New York Then Washington 261, 1032 (1993).
Acknowledgements
This study was supported by the National Natural Science Foundation of China (31661143047, 31470700 and 31270736).
Author information
Authors and Affiliations
Contributions
Yu Y. and Liu J. designed the research. Yang W., Cai Y., Hu L., Wei Q., Chen G., Bai M., and Wu H. performed the research. Yu Y. and Yang W. wrote the paper.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Yang, W., Cai, Y., Hu, L. et al. PhCESA3 silencing inhibits elongation and stimulates radial expansion in petunia. Sci Rep 7, 41471 (2017). https://doi.org/10.1038/srep41471
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep41471
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
